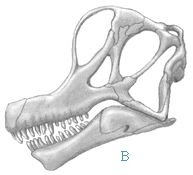
Q: I have heard that the Jack Horner theory about Torosaurus being nothing more than old individuals of Triceratops is getting a lot of pushback. Paleo King, what are your views on the theory, and what does the evidence actually say? A: They are arm-waving. Horner actually has a pattern of doing this, it isn't the first time. Remember the “obligate scavenger T. rex”? Every piece of evidence Bakker, Currie, Weishampel, etc. looked at, Horner basically ignored or dismissed or even denied – even things as basic as eye socket shape – just to preserve his precious theory. Later Horner confessed that he “never liked T. rex anyway” (he was always primarily a duckbill specialist, whose best work was with the
Maiasaura nests) and never took the “100% scavenger idea” seriously, he simply went on TV and made these claims because he wanted to stir debate and knock T. rex down a few pegs. But making an argument you
know is weak simply to stir debate, is the most
unproductive sort of debate there is. Paleo-trolling before there were tumblr and buzzfeed.
You'd think his fans would have learned from that whole fiasco. Most of them don't even know it happened. Score? Hornerite groupies: 0. Horner: 1
As for the “Toroceratops” theory – although Horner (but moreso his former student John Scannella) actually published academic papers and piling up skulls on this new idea, unlike with the scevenging T. rex theory, the line of argument in the papers is hardly any stronger, and the piles of Triceratops and Torosaurus skulls he cites as proof,
do not actually support his claims. The theory basically runs like this: Triceratops and Torosaurus are found in the same rock layers and general region, they look very similar, both have metaplastic bone in their frills indicating bone remodelling, hence “obviously” one must be a growth stage of the other. Except that there's no real conclusive proof for that argument. The fossil evidence in their papers is either circumstantial or not in support of their theory, and some of it is even heavily altered with plaster. The make conclusions about the skulls, which the skulls themselves actually contradict! Same can be said for much of their Pachy-stygi-dracorex theory, which makes a ton of assumptions based on casts, stress fractures, and artifacts of preservation rather than native features of the fossils, as well as for Fowler's "Haplo-Suuwa-Bronto-Diplodocus"
faux-pas, where
downright bizarre fictions like "weighted characters" and "selective parsimony" had to be invented to turn sauropods that are clearly NOT diplodocids into good little juvenile diplodocids (just ignore the fact that
Haplocanthosaurus is also known from non-bifurcated adult neck remains too, right?).
![]() |
| Don't you get all bifurcated on me too now.... |
There's no way to prove that Torosaurus was simply an old adult Triceratops. There are some VERY irreconcilable differences and between the two, and some gaping holes and assumptions in their hypothesis - even if you use ontogeny to "rationalize away" some of the variation. How Jack Horner and his team can hope to bridge these holes in their theory, I honestly have no clue:
1. The changes required are too radical for an animal that is no longer juvenile and has no growing left to do in the postcrania. Such an extreme and late change in facial geometry is unknown in any ceratopsid.
2. Torosaurus is far rarer than Triceratops, too rare to simply be the same animal a few years older. 90% of Hell Creek's large herbivores are Triceratops, Torosaurus forms less than 1%. Some may argue that this is only because few herbivores survived to old age, or that older individuals were more vulnerable to be eaten rather than fossilized – but this is pure speculation, and reason # 4 forces us to discard this idea.
3. Torosaurus has many more epoccipitals than Triceratops - and not only that, their numbers are far more variable in Torosaurus (from 30 to 37), while Triceratops always has exactly 17 (except in elderly skulls where they are reabsorbed). Scannella at SVP claimed that "perhaps Triceratops split their epoccipitals in half to double the number" - something that I can only guess was tongue-in-cheek, since there is no specimen showing evidence for such "stud splits" in either Triceratops, Torosaurus, or any other ceratopsid, much less any dinosaur, period. It sounds even more ridiculous when you realize that Triceratops actually LOST frill studs as it aged, it didn't add new ones. Those near the bottom of the frill tended to disappear first, and in the most mature Triceratops skulls, all of them are gone. To add new frill studs would be a
reversal of the entire Triceratops aging process.
4. Torosaurus has never been found in association with Triceratops.If they were simply aged Triceratops, they would realistically be found in the same herds/locations at least some of the time, especially if they were weaker than young adult Triceratops and needed the protection. This association has
never been known to occur for Torosaurus. You just don't find them at the same dig sites as Triceratops.
5. Different head/body proportions, even discounting the frill. The Milwaukee specimen of Torosaurus
[link] has larger skull (including jugals) than Triceratops, but much smaller postcrania. Doesn't make sense why an adult animal's individual bones would be smaller than those of a "juvenile". Of course Horner doesn't specify if his 30-footers are really juvenile Triceratops or middle-aged adults, he just claims Torosaurus are "old adults". Ok whatever.
6. Torosaurus is actually smaller than Triceratops. As in, the entire body except for the head, is smaller – every bone is smaller than in the most mature Triceratops. How can an “adult” of one species have a smaller body than the “young”, and even smaller limb bones, ribs, and vertebrae? (every postcranial element from Torosaurus indicates an animal 24 ft. long, not 30 ft. as in the most mature Triceratops skeletons! I know humans and some animals lose height as they age due to cartilage contraction, but shrink the femur, the ribs and the humerus? SERIOUSLY? These parts don't even have any metaplastic bone in Torosaurus, nor do they show any proof of bone reabsorption!)
7. Horner and Scannella have never done a histological age analysis on Torosaurus postcraniato actually prove that they are any older than the biggest accepted Triceratops specimens. This would be far easier than guessing age from the skulls, since as mentioned above, postcrania do not typically feature metaplastic bone or remodeling. Not that being older would actually prove that they're old
Triceratops per se, but it would at least remove one
huge logical impediment that stands in the way of Horner's theory being more accepted – on the other hand, a young age for these bones would nuke it.
8. Beak shape in complete skulls is radically different - Triceratops had a strongly hooked, recurved "eagle" beak, while Torosaurus has a much less curved "condor" beak that slopes downwards and forwards, the traditional Chasmosaurine beak design. The sloping “condor” beak is also present in Eotriceratops, which is otherwise far closer to Triceratops proportions than to Torosaurus – putting even more taxonomic space between them.
9. Nasal horn position (and snout/beak length ratio) differs between Torosaurus and triceratops. Torosaurus has shorter post-nasal-horn snout and a longer beak. No ceratopsian is known to radically change beak shape or beak/snout ratios when reaching maturity.
10. Even in very old Triceratops, Torosaurus-like features are extremely rare. The only Triceratops specimen that shows even remotely Torosaurus-like snout proportions and horns is the Torrington skull
[link] , which is a very old adult with reabsorbed frill studs but NO fenestrae in the frill! This may be the most basal Triceratops morph, close to a fork with Torosaurus. Even so, the beak curvature is still not identical to Torosaurus.
11. There appears to be ontogenic variation within Torosaurus itself![link] The Yale specimen, the MOR specimens, the Denver specimen and others all show a great deal of variation in epoccipital reabsorption and horn curvature - indicating that Torosaurus a unique creature that underwent ontogenic changes and growth stages of its own, and not merely a “final phase” of Triceratops.
12. There are other ceratopsids even closer to Triceratops (like Eotriceratops, Nedoceratops, and Ojoceratops)
that don't fit comfortably in any part of Horner's ontogeny sequence. Nedoceratops is the real wild card, as Horner and Scannella claim it as proof of the Trikes aging into Toros - but while it has a few things in common with both,
it fails dismally as a "midlife crisis" stage between the two, since it has a very odd
mix of features that, taken together, make no sense in a Triceratops growth series. They are only found
individually in Triceratops of completely
contradictory age groups, and most of them are not found in Torosaurus at all. Nedoceratops has a very "perky" high horn angle completely inconsistent with the forward curvature of the horns themselves as well as the skull's advanced ontogeny, if one were following Horner's theory
[link] to its logical conclusions. Its horns curve like an old Trike, but their bases are angled up and back like a baby Trike. It beak roughly follows Torosaurus (almost nothing else on its face does) and Eotriceratops, but is shorter than in either, and its frill is essentially a more compact version of Eotriceratops - far shorter and more compact than a Toro frill, yet obviously also older than many Toro frills, judging by its heavily reabsorbed epoccipitals. Its squamosals don't look like anything known in either Triceratops or Torosaurus. Horner and Scannella's nomination of this skull as an ontogenic “transition” from Triceratops-morphs to Torosaurus-morphs is laughable. There isn't a hint of Torosaurus in its short frill length, low epoccipital count, or steep horn angle, and especially not in the squamosal. There is also a second, larger skull that appears to be Nedoceratops - CMN 8862, which was once labeled "Triceratops albertensis". It has the same "perky" horns and the same short, upcurved, very un-Toro-like squamosal. Clearly this animal had a different ontogeny pattern as it grew than either Triceratops or Torosaurus. It likely branched off from the family tree sometime after Torosaurus and before Eotriceratops.
13. Fake plaster fillers are misleading. The beak of the YPM Torosaurus skull, as well as the horn tips and rear frill of some Torosaurus skulls and most of the Milwaukee specimen skull have been incorrectly reconstructed to look like Triceratops. Also the MOR skulls have a huge nasal boss in place of a horn, which is not consistent with anything seen in Triceratops, least of all the beak structure. It's actually a bit shocking how the most commonly pictured Torosaurus skulls have FAKE BEAKS and FAKE skull fullers in general that are modeled on Triceratops skulls, rather than more complete Torosaurus skulls - GetAwayTrike faithfully reproduces both the complete skulls and the fragmentary ones with fake Triceratops-mimic plaster fillers (recurved eagle beaks, short snouts, etc.),
the differences are often extreme. Of course some of this error was probably due to lack of access to all the Torosaurus material, and earlier date of discovery/preparation with some specimens, but still... most of what you may THINK are correct Torosaurus orbital, snout, and beak features, are FAKE. Those that have a mostly complete beak, like ANSP 15192 and the far larger MOR 981, show a VERY different beak structure than in mature Triceratops.
14. The degree of cranial variation between Triceratops and Torosaurus is greater than that between many closely related modern bird, reptile, and mammal genera. Take antelopes as an example: ignoring the keratin horn sheaths, the actual skeletons and skulls that can fossilize are VERY hard to tell apart. There is hardly any cranial variation comparable to that between Triceratops and Torosaurus. Are we then to conclude that the Gemsbok is simply an immature Eland, or that the Springbok is an ontogenic stage of the Thompson's Gazelle? A better case could be made for using Horner's lumpery on these, than on Torosaurus, even though we clearly know these animals are not growth stages of each other. Impalas don't turn into Heartebeest, even though their bodies are basically the same design on different scales! There's a lot of diversity even in unhealthy human-damaged ecosystems. So even if the Maastrichtian faunas of the Rockies were doing badly in terms of diversity, it's doubtful that triceratops was the ONLY horned dinosaur there. The presence of Ojoceratops, Tatankaceratops, Eotriceratops, all of which are more Triceratops-like than Toro, further confirms this.
15. The raw morphometric data does not support lumping them. Farke, et. al. (2013) determined the changes required to "age" a Triceratops into a Torosaurus to be UNPRECEDENTED among ceratopsids, requiring
addition of epoccipitals (frill studs), reversion of bone texture
from adult to immature back to adult, and unusually late growth of holes in the frill. The Torosaurus specimens cluster together,
separate from the Triceratops cluster on the morphometric plot.
16. Torosaurus has its own immature specimens. These, such as the ANSP skull, have a shorter and more upcurved frill that had yet to fully flatten out, and their beak shape, horn angle, and fenestrated frills are still clearly distinct from Triceratops of the same growth stage (or
any growth stage for that matter). It's not so easy to claim that Torosaurus is the mature form of Triceratops, when it has its own juvenile specimens that are clearly NOT Triceratops. They even have high frill stud counts, just like the Torosaurus adults. Clearly these animals did not closely resemble Triceratops, even when less than half-grown.
17. Large parts of Trike's and Toro's ranges do not overlap. Juvenile Torosaurus were found in Big Bend National Park, Texas, which is conspicuously devoid of Triceratops material - though Ojoceratops is present in nearby New Mexico – a compact, short-faced and short-frilled animal, almost as different from Torosaurus as it is possible for a derived chasmosaurine to be. Ceratopsid faunas, it seems, were far more diverse than the plain one-genus badlands Horner would prefer them to be.
18. Many large Torosaurus are less mature than the most mature Triceratops. Most of the Torosaurus skulls, including the largest ones, actually appear to be ontogenically LESS mature than the largest Triceratops skulls. This is true both in terms of epoccipital reabsorption and the amount of metaplastic bone. The MOR skulls in particular are gigantic, but clearly immature, having young, well-defined epoccipitals/epipareitals and relatively small fenestrae, which may mark them out as a unique new species within Torosaurus itself.
19. Torosaurus horns typically look more like teenage Triceratops than mature ones. The most complete Torosaurus skulls all have relatively slender and typically straight horns, and some have a slight double curve - not the thick robust forward-curving horns of mature Triceratops. In fact the closest thing to a Torosaurus brow horn among most Triceratops is adolescents or young adults of Triceratops which have barely attained the double curve stage:
[link] let alone the strong forward curve stage of mature Triceratops horridus and prorsus:
[link][link] In order to actually turn the most mature Trikes into Toros, you would have to actually reverse the changes in their horns - undo the mature forward curve, re-lengthen and re-straighten them and in some cases even re-add the double curve found in younger Trikes - all while the postcrania mysteriously shrink by 25%! What a completely unnatural and pointless waste of metabolic processes and resources. And it doesn't happen in ANY other ceratopsid known to man.
So did mature Triceratops just straighten out their horns a SECOND time and make them longer and slimmer after having already absorbed the tips and thickened the bases
[link] ,
reversing much of the normal Triceratops aging process, all to become Torosaurus? I doubt it. There's no "Benjamin Buttonceratops" in Hell Creek, we're simply looking at two genera where the ontogeny changes worked differently.
20. Mature Triceratops specimens actually appear to be shortening the frill, not lengthening it. This goes along with the fact that they were reabsorbing and in some cases losing frill studs, not growing extra ones. The frills of many old Triceratops skulls are, if anything, receding. This is especially the case with T. prorsus, but it appears in some old T. horridus skulls as well. MOR 004 (prorsus), SMNH P1163.4 (prorsus), TCM 2001.93.1 (horridus), and several others, have the forward-curved horns and reabsorbed tips of old individuals, but the frill is moving in anything
but a Torosaurus-like direction. If anything, it's proportionally shorter and more compact than in the less mature large Triceratops specimens with double-curved horns. Once again, just like with the horns and frill studs, lengthening the frill or growing new frill in these mature skulls would actually be a
reversal of the normal Triceratops aging process up to that point. Some T. horridus individuals like UCMP 113697 do have a longer frill, but still lack the fenestrae of Torosaurus, and the epoccipitals are long gone, with no hint of them re-emerging and doubling in number to "become" a Torosaurus frill edge. You can check out some of these skulls
here; the original drawing is by GetAwayTrike; please do note that this diagram isn't strictly Triceratops skulls, it also throws in the type skull of Nedoceratops and a referrable skull from the Scollard formation (which predates Hell Creek and true Triceratops), albeit with the missing snout restored like a Triceratops, and also includes Eotriceratops from the Horseshoe Canyon formation and Ojoceratops from Ojo Alamo.
![]() |
| Torosaurus and Triceratops adults and juveniles to the same scale (the adult Toro's beak is restored with the tip pf the juvenile's beak scaled up, as the original was eroded - other large Toro skulls have the same "condor" beak tip). Trike and Toro look really different as youngsters and even up even MORE different as adults. The frill is basically doing opposite things in both genera as it matures (extending and flattening vs. getting more upcurved and turning in on itself). Also note that even young Torosaurus had fenestrae in the frill, and that as adults, Torosaurus overall has a much larger frill than Triceratops, but a smaller face not counting the frill. |
The big picture beyond Horner and "Toroceratops": Why the "hyper-lumper" approach is probably WAY off-base Considering how much we know about both mammalian and avian biodiversity in recent ages like the Miocene, Pliocene and Pleistocene, and also given how much less terrestrial fossil material of any sort inevitably survives over time from older epochs like the Maastrichtian (and even less from earlier times), it may actually make the most sense to say that dinosaur faunas were MORE diverse and had MORE genera and species than we can ever possibly know - likely far richer and more diverse than mammalian faunas today, perhaps even more so than Pliocene and Pleistocene faunas. We just don't have as complete a fossil record when you go back into the mesozoic. Even so, recent discoveries have more than doubled the number of maastrichtian ceratopsids known. We're well past the point of "only Trike and Toro" in the US/Canada Maastrichtian time horizon, and any proper morphometric character analysis will show that there are a number of evolutionary steps between Torosaurus and Triceratops, which form their own unique genera.
Nonetheless, the view is still far from complete. Some species have probably never had a single individual get fossilized, which isn't all that strange when warm-blooded species sometimes last for less than a million years. Or we may only get one skull from an entire genus, because geological processes may have jumbled the rock layers so much that we will never have access to more than that, either due to their destruction in these processes, or their being buried in inaccessible depths, with no surface hints of their presence.
Now if that one skull happens to have some similarities with an already known genus, say, Triceratops (???), people will be tempted to gloss over the differences and lump it into Triceratops, even if some parts of it don't quite fit anywhere within known Triceratops populations and growth stages. But then, is it
really an odd growth stage or an abnormal individual of Triceratops, or simply something else we don't properly understand yet? Then, when you actually find more growth stages and skulls of the new animal that show it's a unique genus with its own ontogeny pattern (like we now have with Torosaurus), what can you honestly say for a person who persists in denying its status as a separate genus or holding the clear differences to be one-off aberrations of no account? This might have been plausible when there were only one or two Torosaurus specimens known to science, but now there are over thirteen of them from different growth stages, and possibly comprising three different species (T. latus, T. utahensis, and "T. magnus", i.e. the MOR skulls).
Insistence on lumping two genera together "because they both have metaplastic bone" really is pointless. As it is, metaplastic bone is not exclusive to any one genus or growth stage, and we're already over-lumping extinct specimens
based on arbitrary standards that would make no sense to a biologist studying living animals. There is less difference between the skull morphs of cheetahs and jaguars than you get between Trike and Toro - but nobody is proposing to lump cheetahs and jaguars into the same genus - DNA cladistics finds
no less than FOUR other cat genera separating them.
The same phenomenon of morphological similarity "masking" generic diversity is found all over the place, whether in Birds of Paradise or in the host of antelope genera that cannot hybridize but look nearly identical when you get rid of the keratin horn sheaths (which would not fossilize). Of course you can achieve high morphological diversity in skeletons without genetic diversity, but aside from artificial selection by humans (as in the case of dog breeds) and a few extreme cases of sexual dimorphism, it's extremely uncommon in any vertebrates. We don't have DNA from dinosaurs, but if morphometrics are any clue, dinosaur paleontologists are lumping at a generic level,
far more than they would be if such DNA existed. I am not suggesting we go back to having 16 species of Triceratops like in the 1950s (some of which were nearly identical to each other, and some of which were actually other genera like Nedoceratops). But on the generic level, things are definitely overlumped - something that even a good non-DNA-dependent analysis like Tschopp et. al.'s diplodocoid paper can expose very well.
If anything, the mainstream view of dinosaurs is actually already overlumped, even without Horner and Scannella's antics. As cladistic science gets more precise and uses more and better characters (and weeds out coding errors better), this is already becoming more apparent. Giraffatitan and Lusotitan are no longer part of Brachiosaurus. Galeamopus is no longer in Diplodocus. Traukutitan is no longer part of Epachthosaurus. Isisaurus is no longer in Titanosaurus. Brontosaurus - all 4 or 5 species of it - is no longer in the Apatosaurus wastebasket. It's an open secret that Mamenchisaurus and Omeisaurus between them currently contain around 10 other genera that should be spun off. And it should be obvious that not every ceratopsid with metaplastic bone is a growth stage of Triceratops. And of course, that Kosmoceratops is NOT a juvenile Utahceratops (what happened in Vegas... lol).
![]() |
| Dude, we don't even have the same frill stud arrangement, never mind number... |
Do we really believe that metaplastic bone only exists in one species, or in just one ontogenic stage for any given species? Heck, even if Trike and Toro had identical ontogenic changes in horn and frill shape as they matured, or even if one's ontogeny pattern
appeared to neatly transition into the other's (they don't, not even close), the fact that they both have metaplastic bone throughout multiple growth stages proves NOTHING conclusive in favor of lumping the two together. At best,
even if the ontogeny changes matched or appeared to dovetail, and
even if we didn't have inconvenient things like the ANSP skull or the Big Bend Toro juveniles to sour the deal for the Hornerites, it's still possible that these could be
no more than two related genera with similar growth patterns. Even very old ceratopsids have metaplastic bone in their skulls, it's not proof of immaturity or an upcoming radical change in head shape - and even then, you could probably make a far better (though still wrong) morphological case for lumping Eotriceratops or Ojoceratops into Triceratops (despite the geographic and time discrepancy)
long before you get to Torosaurus.
In many cases the metaplastic bone may have nothing to do with age, and far more to do with rapidly healing injuries and getting rid of infection (bone cells that naturally die rapidly and are replaced by new ones from below in a constant conveyor-belt cycle, are far less susceptible to infection - and we know these animals, with their vein-engorged frill bones, were just as susceptible to injury and possible infection from each other's horns as from a tyrannosaur bite to the face). Many of the irregular holes in "pathological" ceratopsid skulls of various genera and ages are bordered by metaplastic bone, did they ever stop and think what the connection was there? There's definitely a paper in that. So there are a lot more plausible alternate reasons for metaplasia that Horner and co.
don't even address. Is their "Toroceratops" theory still possible? Sure, but the amount of contortion (both osteological and rhetorical) required
merely to make it work also violates Occam's razor repeatedly. You'd literally have to ignore
everything they didn't figure in their papers (even when they mentioned an inconvenient specimen in passing) and also ignore how ontogeny works in
every other well-represented ceratopsid.
![]() |
| Get Away Trike! This is Toro time! Note that the T. utahensis material is smaller than most "teenage" Triceratops let a lone the older ones. The two MOR skulls have nasal horns and beaks unlike ANYTHING found in Triceratops - and they were still growing. |
You want more details on Horner's lumpermania with Pachycephalosaurs too? Find it
here and
here.